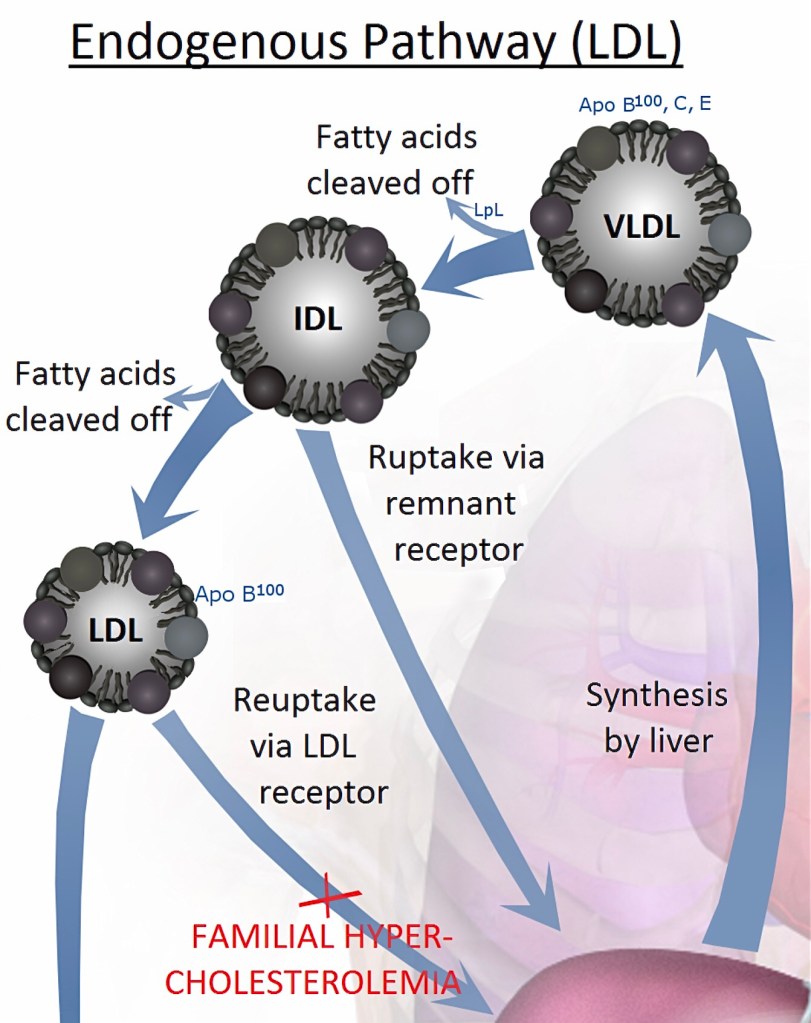
LDL är ett lipoprotein som börjar som VLDL i levern för att sedan gradvis krympa till IDL och vidare. LDL tillhörde de 50% av IDL som i slutändan av sin utveckling bara behöll kännetecknet ApoB-100 och därför slipper förbi leverns utrensningsmekanism. Se föregående inlägg.
Varje LDL transporterar 3000-6000 triglycerider (fettmolekyler), obundna kolesterolmolekyler, kolesterylestrar samt fosfolipider som ingår i dess membran (se föregående delar för ytterligare information). Vid brist på LDL faller ansvaret för framställning av kolesterol på de enskilda cellerna, vilket de inte alltid klarar av. Celler som använder LDL (med ApoB-100) som energileverantör kan inte identifiera och hämta sitt behov från kylomikroner med ApoB-48 som sitt “ID”.
I varje LDL finns bland andra fetter även linoleate, innehållande omega-6-fettsyran linolsyra. Fleromättade fettsyror har två eller flera dubbelbindningar i sina kolkedjor, alla känsliga för angrepp från fria radikaler, de oxideras lätt.
Celler har LDL-receptorer som går upp till cellmembranet och flyttar sig till område med urgröpningar (clathrin-coated pits) på cellmembranets utsida. Där stannar den och ”fiskar” efter passerande lipoproteiner med ApoB-100 där den kan hämta mer. När den ”får napp” sluter sig urgröpningen, dras i form av en vesikel in i cellen (endocytos) där den öppnar sig och lämnar av sin fångst. Därefter kan LDL-receptorn återgå till cellytan och upprepa processen. Om deras insats inte längre krävs går de till cellens lysosomer för återvinning av sina beståndsdelar. De flesta av våra celler kan dessutom själva framställ kolesterol för eget bruk.
När koncentrationen av kolesterol inne i cellen är tillräcklig deaktiveras enzymet HMG-CoA reduktas vilket dämpar den interna kolesterolproduktionen. På så sätt avlastas cellen från en del av arbetet om man äter mat som innehåller kolesterol. Samtidigt minskar nyproduktionen av LDL-receptorer och eventuellt överskott av kolesterol förestras med fettsyror till en kolesterylestrar, bättre anpassade för lagring.
Statiner är HMG-CoA-reduktashämmare som verkar generellt i kroppen, inte bara i de celler där behovet av nytt kolesterol är tillräckligt.
LDL finns av flera subtyper varav A beskrivs som ”stora och fluffiga” (large and fluffy) B som ”små och täta” (small and dense). B-typen (pattern B) utpekas som den skadliga varianten då deras storlek gör att de lättare fastnar i kärlväggarna. I själva verket finns 7 urskiljbara subtyper där de två av de tidiga i förloppet är A-typen och de övriga 5 tillhör B-typen. Bilden nedan redovisar inte subtyperna 6 och 7.
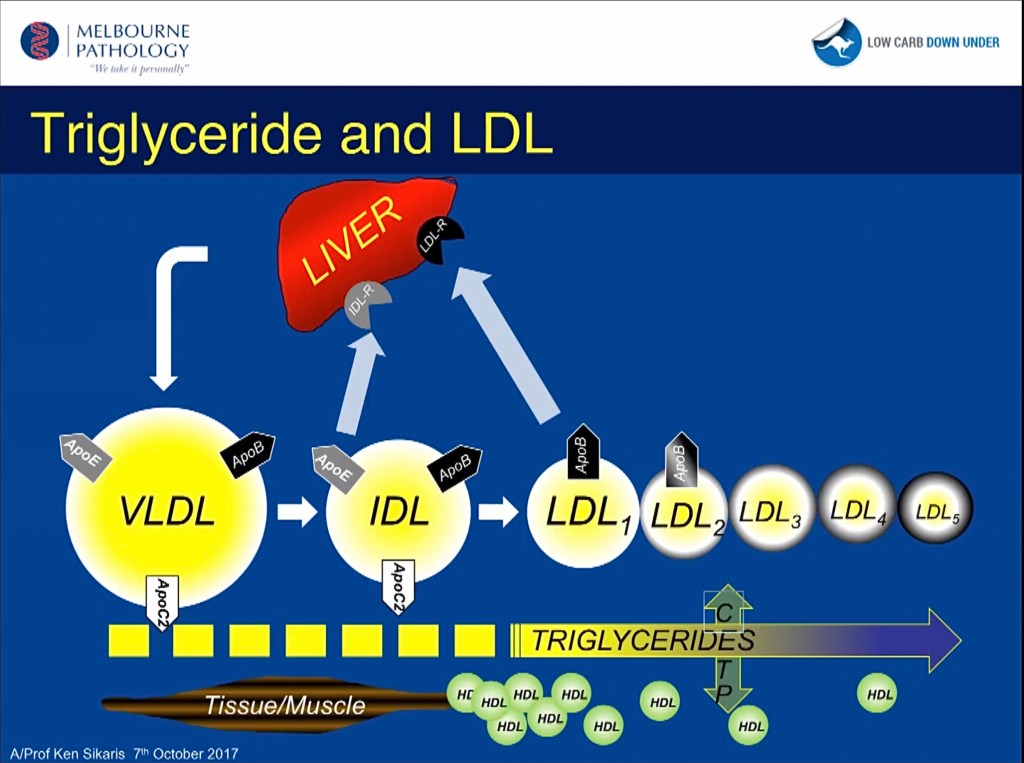
Analyser av labbvärden har visat att höga VLDL (det lipoprotein som levern tillverkar och använder för att exportera sitt triglyceridbidrag) är associerade med mer LDLsd (small dense), B-typen.
Vid vanliga blodprov mäter man totalkolesterol, HDL (nämnts tidigare men ännu inte berört) samt triglycerider, TG (fettmolekyler) i lipoproteinet VLDL. Ur dessa värden gör man uppskattningar ”mellan tummen och pekfingret” enligt Friedewalds formel:
LDL (alla 7 fraktioner tillsammans) ≈ (ungefär) Totalkolesterol – HDL – 0,45 * TG där alla värden anges i mmol/L.
Formeln är på inga sätt exakt och förutsätter en föregående nattfasta på mer än 12 timmar för att TG-värdet i huvudsak ska återspegla fett som för ögonblicket levereras ut från levern. Dessutom förutsätter den antaganden om genomsnittsinnehållet i de olika lipoproteinerna, något som kan variera rejält. Men metoden är förhållandevis billig och tjänar väl att skrämmas med.
En betydligt bättre variant är att räkna antalet ApoB-100 (från LDL) och ApoA1 (från HDL). Kvoten ApoB-100/ApoA1 (Apo-kvoten) bör vara under 0,9 för män och under 0,8 för kvinnor för att betraktas som OK, lägre anses bättre. Vill din läkare att du ska ta kolesterolsänkande mediciner så be åtminstone först om en Apo-kvot som stöd för åsikten.
Nästa inlägg innehåller mer om LDL.